
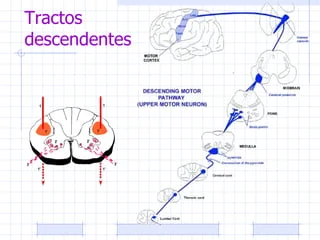
A grandes rasgos, cuatro son las vías descendentes o motoras (corticoespinal, retículoespinal, rubroespinal y vestíbuloespinal) sobre las cuales, nuestro Sistema Nervioso Central manda sus señales al resto de la periferia, para ejecutar a su «antojo», las órdenes activas sobre las cuales nos dirigimos para cambiar nuestro mundo, tanto para ser un medio como un fin, el movimiento.
Y es que las funciones organizativas de cada una de ellas pueden ser interesantes a la hora de rehabilitar una disfunción en el movimiento, ocasionado por una lesión de una de las vías que implican una clínica específica ya investigada. Muchas veces, por la situación concreta en la que se ha desarrollado un ictus o un Daño Cerebral Adquirido (DCA), siendo ésta una zona de paso de muchas vías descendentes (como por ejemplo, la muy sufrida cápsula interna, zona de paso de varias vías descendentes), cabe destacar que existe una variabilidad clínica muy relevante, ya que la afectación al ser una «zona de paso», atribuye daño de manera aleatoria y muy poco presentada en patrones.
Este interesante artículo (1), donde desarrollan el estudio específico de la organización motora de la mano de los macacos, a través de la provocación de una lesión neurológica de un tracto descendente, concretamente mediante lesiones quirúrgicas bilaterales del tracto corticoespinal (el más grande y más importante de las vías motoras), examinaron cuál era la función principal de éste. Tras dicha lesión, inmediatamente después de la lesión, los animales mostraron una parálisis flácida, pero lo interesante viene después. En los días siguientes recuperaron considerablemente la función motora, de tal manera que podían trepar y correr alrededor de sus jaulas casi con total normalidad. Por contraste con esta función locomotora recuperada, los movimientos de motricidad fina de la mano, permanecieron muy deteriorados, y nunca lograron recuperarlos, siendo éstos el sello distintivo de la destreza manual de los primates.
Para investigar más a fondo cuál de las estructuras permiten la recuperación de la función de la mano en ausencia del tracto corticoespinal, Lawrence & Kuypers (1968b) sometieron a los animales recuperados a lesiones quirúrgicas más selectivas de las vías motoras restantes. Cortaron las vías laterales del tronco cerebral (que comprenden principalmente el tracto rubrospinal) y éstas dieron como resultado una pérdida de agarre con la mano, que nunca se recuperó; los movimientos locomotores brutos (los contrarios a la motricidad fina) estaban relativamente poco afectados. Por el contrario, el corte de el sistema descendente medial del tronco cerebral (principalmente reticuloespinal y vestibuloespinal) produjeron un deterioro severo de los movimientos brutos, pero los animales se mantuvieron capaces de agarrar la comida si se les colocaba cerca de la mano.
Sacando una primera conclusión, la motricidad fina de la mano tiene como componentes más importantes el tracto corticoespinal (Limón, 1993; Schieber, 2004 cf. Schieber, 2011)(movimientos de los dedos independientes y selectivos) así como el rubroespinal (Sasaki et al. 2004)(inervando musculatura más distal).
Por otro lado, el papel del tracto reticuloespinal se ha estudiado intensamente en la marcha, la actuación de los ajustes posturales y en los alcances, verificando su implicación en éstos pero no de manera exclusiva, ya que están coordinados con las salidas de los tractos corticoespinales y rubrosespinales.
Debido a que los conceptos actuales enfatizan el papel de la estimulación reticuloespinal a la hora de realizar los alcances y los movimientos gruesos del aparato locomotor, los estudios que tratan de asignar las salidas de la formación reticular en primates generalmente han ignorado los músculos que actúan sobre los dígitos (Davidson & Buford, 2004, 2006; Davidson et al., 2007). Como comentábamos anteriormente, a los macacos que se les lesionaba el tracto reticuloespinal, podían subir alrededor de sus jaulas, incluyendo el agarre de los barrotes de la jaula de tal manera, que podían soportar su peso corporal total. Otros estudios confirman que la formación reticular, al ser estimulada, podría provocar la actividad en los músculos que actúan alrededor de la muñeca. Además, el reflejo de sobresalto acústico – que es más probable que sea mediada a través del tracto reticuloespinal – puede producir la activación de los músculos intrínsecos de la mano cuando se facilita de forma anormal en los pacientes con hipereflexia (Brown et al 1991c.). Por último, Ziemann et al. (1999) informaron que la estimulación magnética transcraneal sobre la corteza motora primaria en sujetos humanos podría provocar respuestas en músculos de la mano ipsilateral. Las características de estas respuestas sugirieron que estaban mediadas a través de una vía del tronco cerebral (probablemente la reticuloespinal), activada a su vez por las proyecciones corticoreticulares.
Por tanto, en el momento que observemos una presentación clínica de una ausencia de control motor de la mano, y en referencia a la bibliografía expuesta, sería interesante intervenir en la estimulación de los tractos descendentes, donde por ejemplo, ante una muñeca que el paciente no puede estabilizar, el trabajo a través de ejercicios de enderezamiento, alcances o marcha, son interesantes para la búsqueda de esa vía reticuloespinal estimulada de por si, para elaborar un trabajo específico de ese tracto y su implicación en la muñeca. Lo mismo podemos observar en otro tipo de paciente, con presentación clínica de pérdida de movilidad selectiva en los dedos, donde un trabajo del tracto corticoespinal así como el rubroespinal, son más que interesantes para poder llegar a estimular esos dedos, como por ejemplo, trabajar el orbicular de los ojos junto con el de los labios (en una actividad que implique también el agarre de dedos, como pintarse los labios), para al menos estimular éstas vías descendentes.
Bibliografía:
(1) , & ( 1960 ). Descending projections to spinal motor and sensory cell groups in the monkey: cortex versus subcortex . Science 132 , 38 – 40 .
(2) & ( 1968a ) The functional organization of the motor system in the monkey. I. The effects of bilateral pyramidal lesions . Brain 91 , 1 – 14 .
(3) Baker SN. The primate reticulospinal tract, hand function and functional recovery. J Physiol 2011 Dec 1;589(Pt 23):5603-5612.
, & ( 1986 ). Corticospinal facilitation of hand muscles during voluntary movement in the conscious monkey . J Physiol 381 , 497 – 527 .
( 2008 ). Descending pathways in motor control . Annu Rev Neurosci 31 , 195 – 218 .
, , , , , , & ( 2004 ). Dexterous finger movements in primate without monosynaptic corticomotoneuronal excitation . J Neurophysiol 92 , 3142 – 3147 .
(7) , , , & ( 1991a ) The effect of posture on the normal and pathological auditory startle reflex. J Neurol Neurosurg Psychiatry 54 , 892 – 897 .
(8) , , , , , , & ( 1999 ). Dissociation of the pathways mediating ipsilateral and contralateral motor-evoked potentials in human hand and arm muscles . J Physiol 518 , 895 – 906 .


